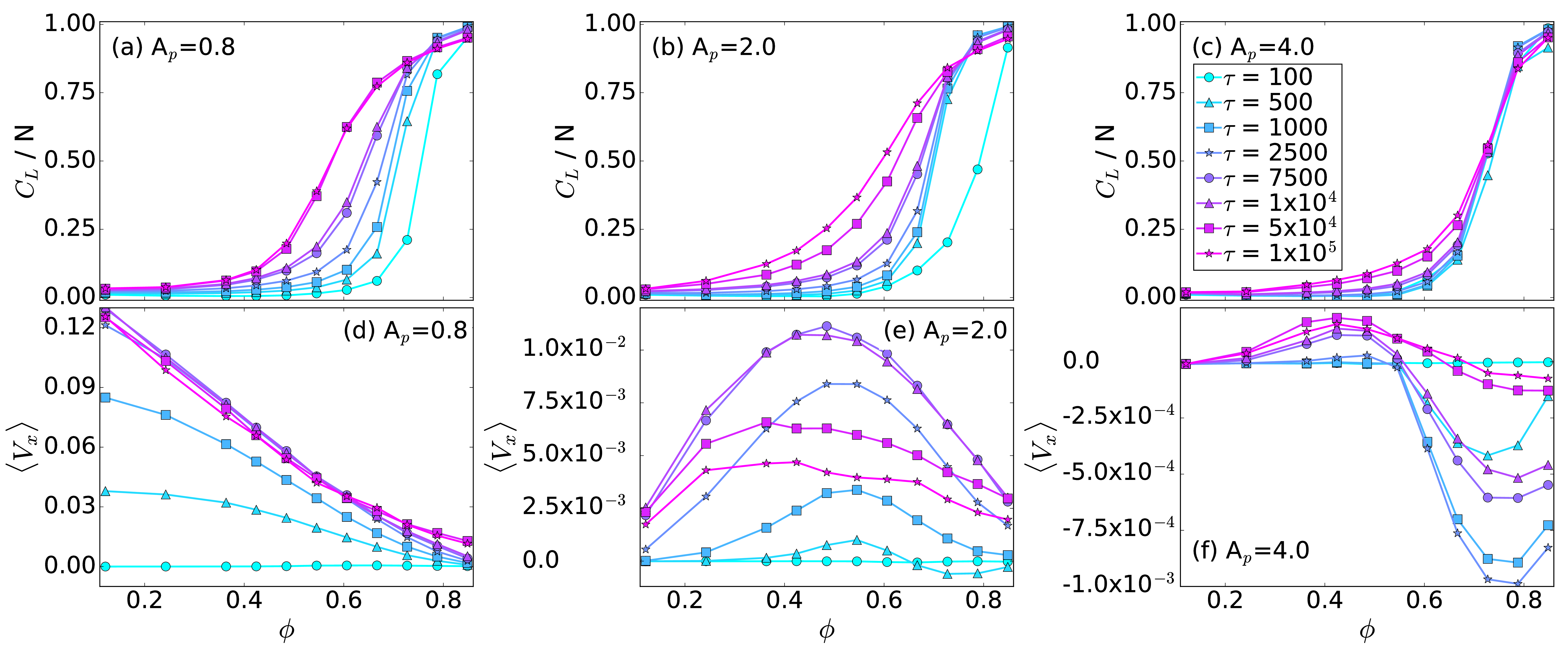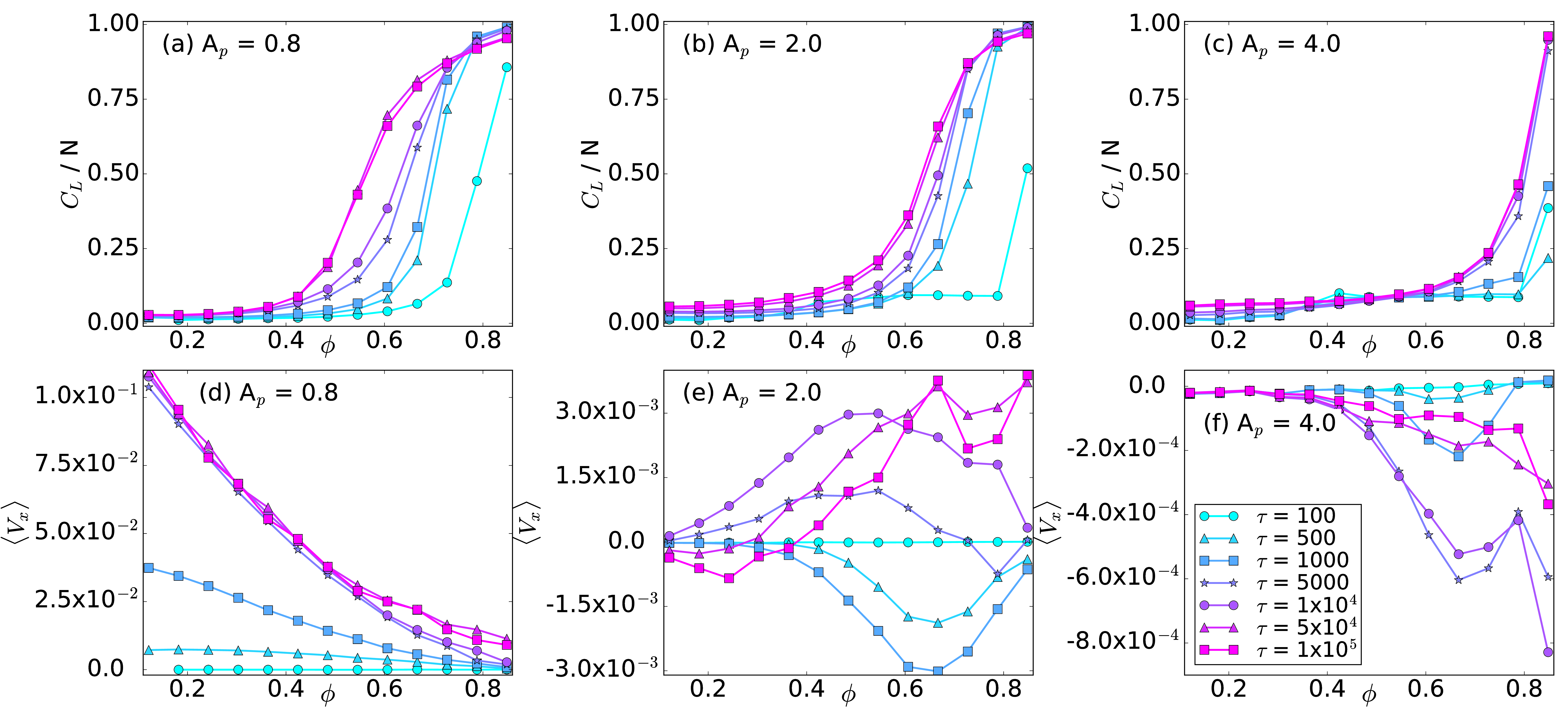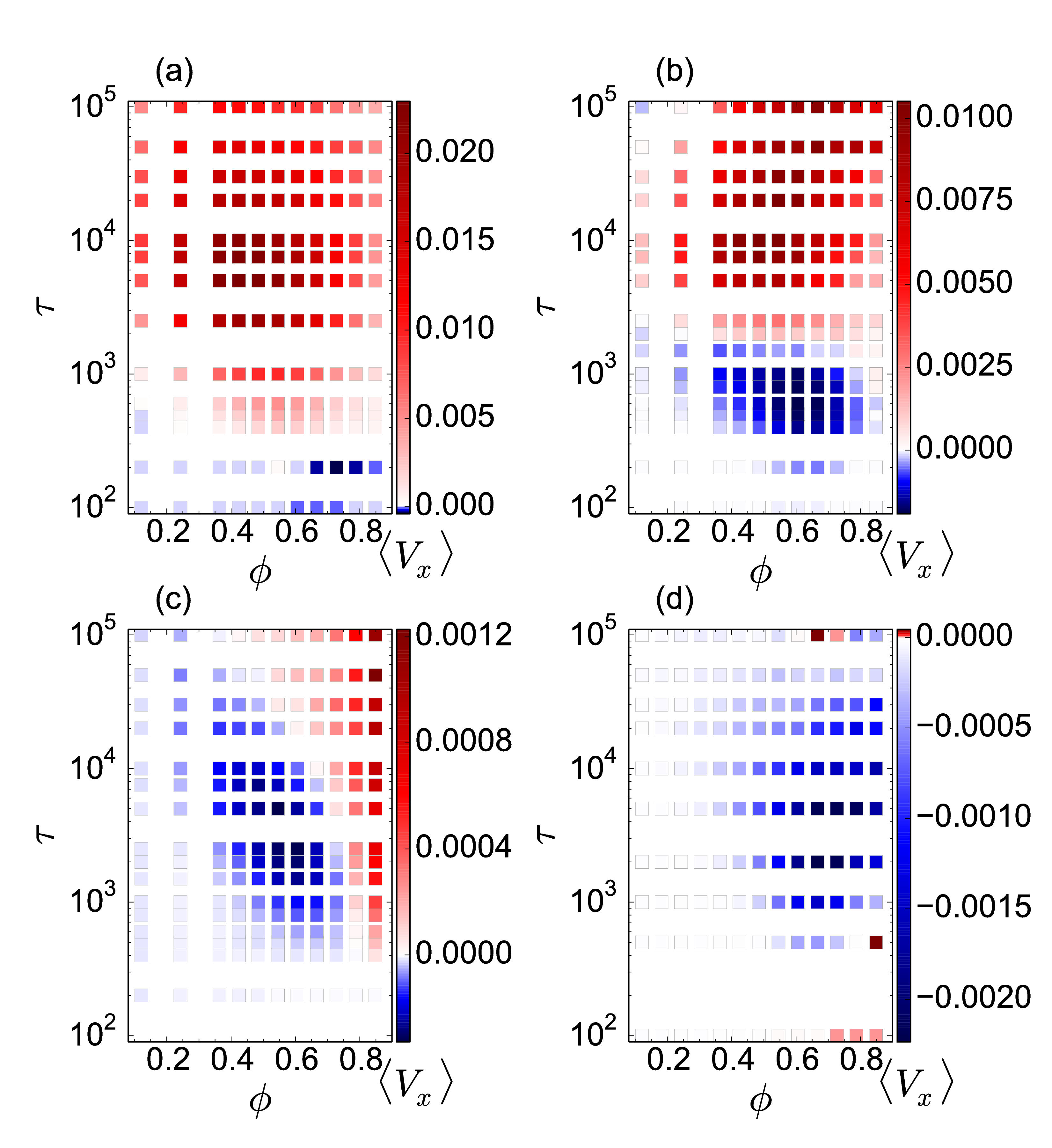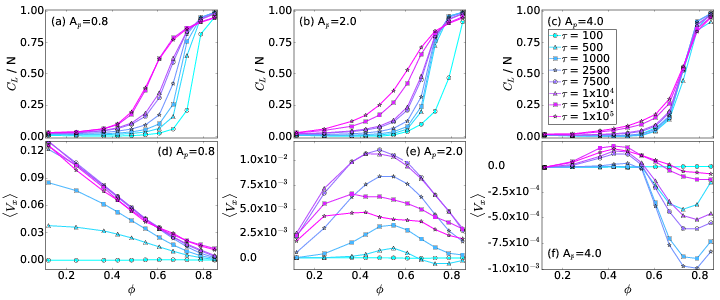
Fig. 2: (a,b,c) The average cluster size CL/N vs particle density ϕ.
(d,e,f) The average particle velocity 〈Vx 〉 vs ϕ.
The color code, shown in panel (f), indicates different
run times
τ = 100, 500, 1000, 2500, 7500,
1×104, 5×104, and 1×105.
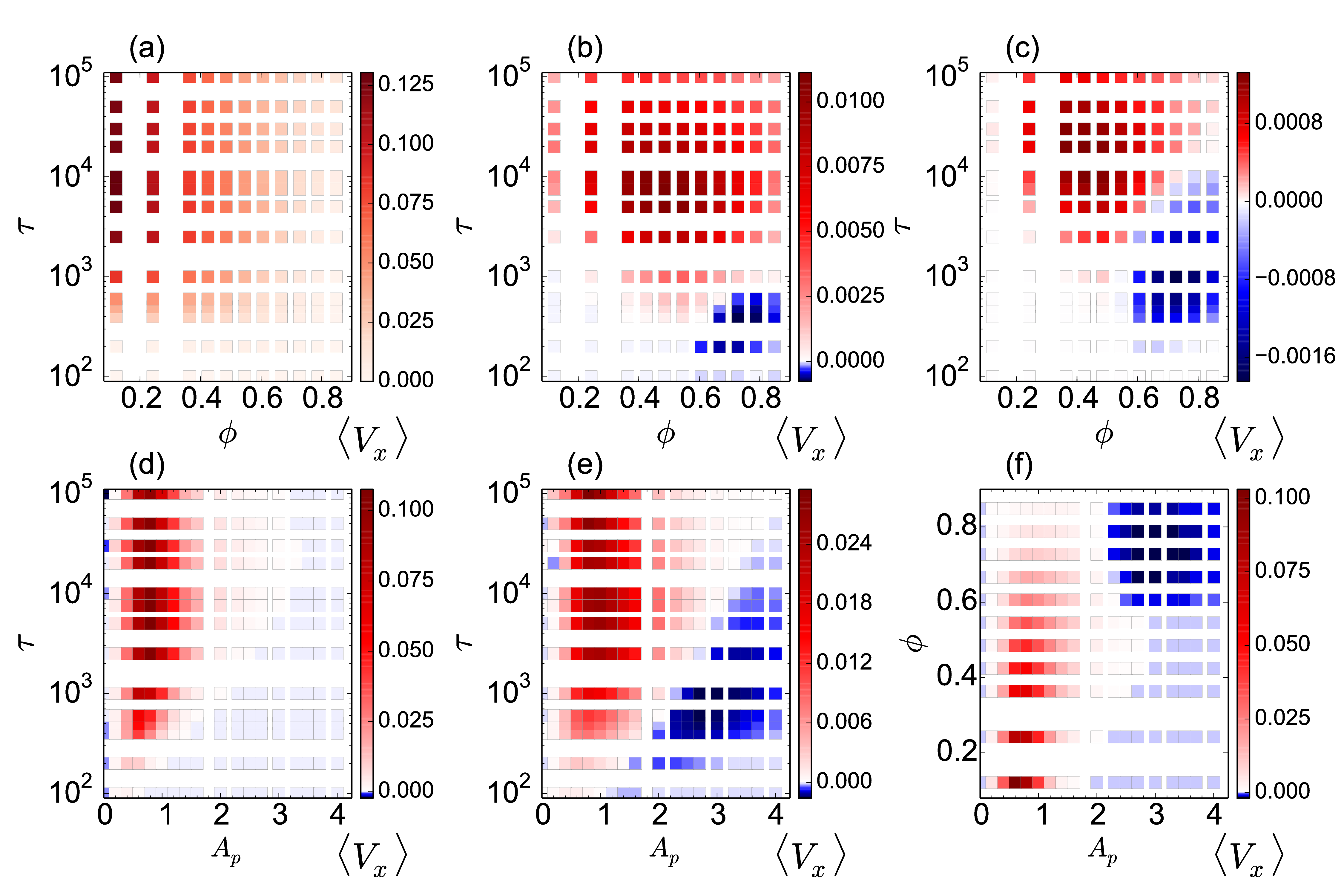
(a,d) At Ap = 0.8,
there is a normal ratchet effect with
a transport flux that
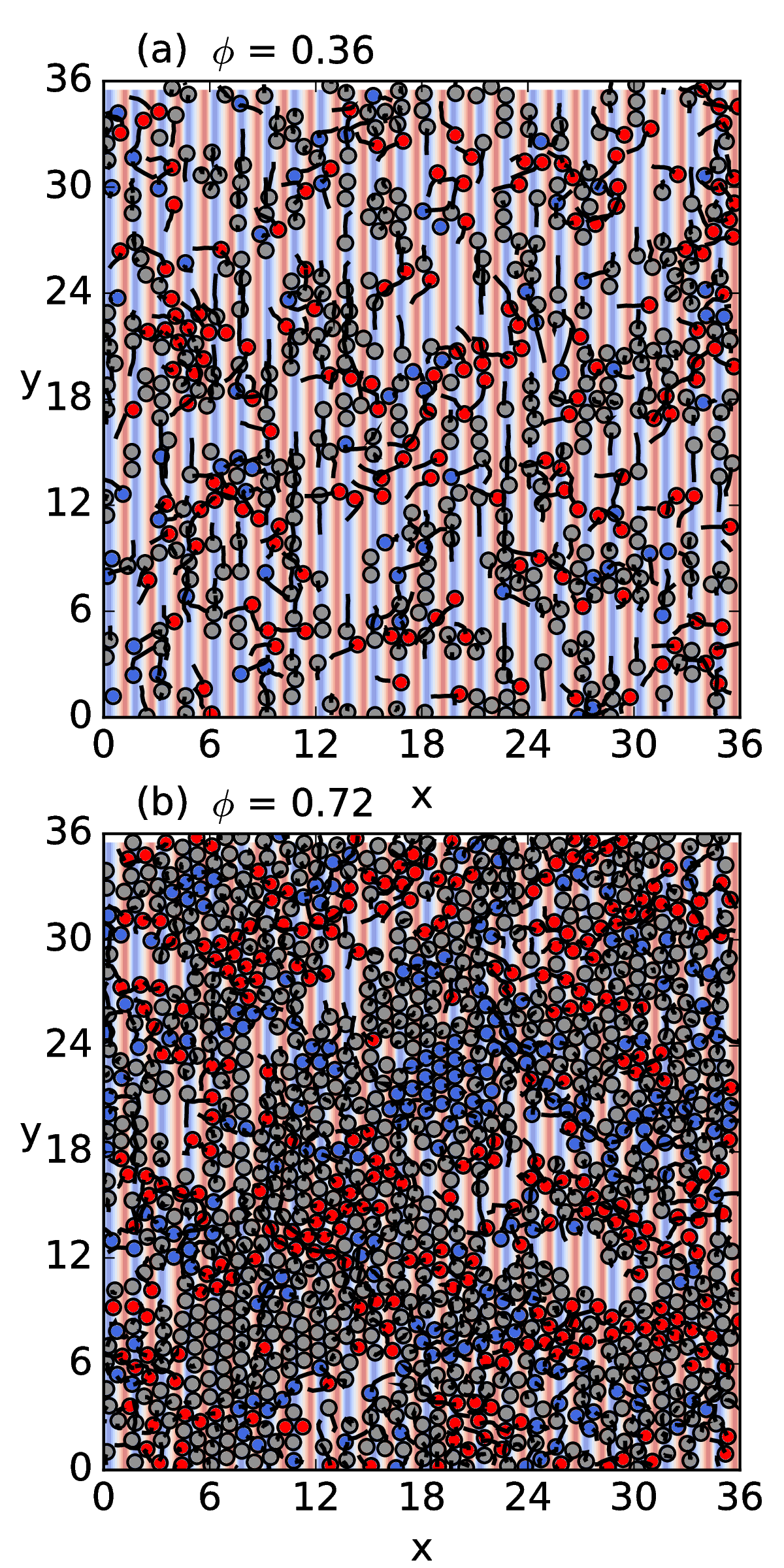
decreases with increasing ϕ.
(b,e) At Ap = 2.0,
the
transport flux of the normal ratchet effect is nonmonotonic,
so that the ratcheting is optimized for a midrange value of ϕ.
(c,f) At Ap = 4.0,
there is a crossover from a normal to a reverse
ratchet effect with increasing ϕ.
|